Регуляция сна и бодрствования. Есть ли у циркадианных ритмов генетический компонент

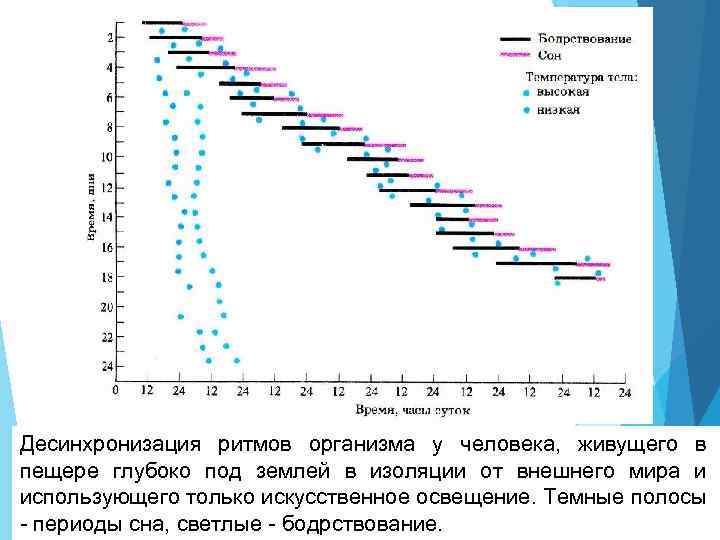 Десинхронизация ритмов организма у человека, живущего в пещере глубоко под землей в изоляции от внешнего мира и использующего только искусственное освещение. Темные полосы - периоды сна, светлые - бодрствование.
Десинхронизация ритмов организма у человека, живущего в пещере глубоко под землей в изоляции от внешнего мира и использующего только искусственное освещение. Темные полосы - периоды сна, светлые - бодрствование.
 Супрахиазматическое ядро расположено над зрительным перекрестом в основании гипоталамуса. Оно получает вход от зрительной системы и отвечает за восприятие светлого времени суток как времени бодрствования, а также поддерживает суточный ритм.
Супрахиазматическое ядро расположено над зрительным перекрестом в основании гипоталамуса. Оно получает вход от зрительной системы и отвечает за восприятие светлого времени суток как времени бодрствования, а также поддерживает суточный ритм.
 Бодрствование - состояние психики, характеризующееся достаточно высоким уровнем электрической активности мозга, свойственным активному взаимодействию индивида с внешним миром. В поддержании состояния бодрствования важнейшую роль играет ретикулярная формация среднего мозга, от нейронов которой восходящие влияния идут к неспецифическим ядрам таламуса, а от них ко всем зонам коры больших полушарий. Бодрствование (обычное состояние не спящего человека) образует поле всевозможных сочетаний функций сознания - от состояния спокойного бодрствования через активное, напряженное бодрствование до выраженных аффектов.
Бодрствование - состояние психики, характеризующееся достаточно высоким уровнем электрической активности мозга, свойственным активному взаимодействию индивида с внешним миром. В поддержании состояния бодрствования важнейшую роль играет ретикулярная формация среднего мозга, от нейронов которой восходящие влияния идут к неспецифическим ядрам таламуса, а от них ко всем зонам коры больших полушарий. Бодрствование (обычное состояние не спящего человека) образует поле всевозможных сочетаний функций сознания - от состояния спокойного бодрствования через активное, напряженное бодрствование до выраженных аффектов.
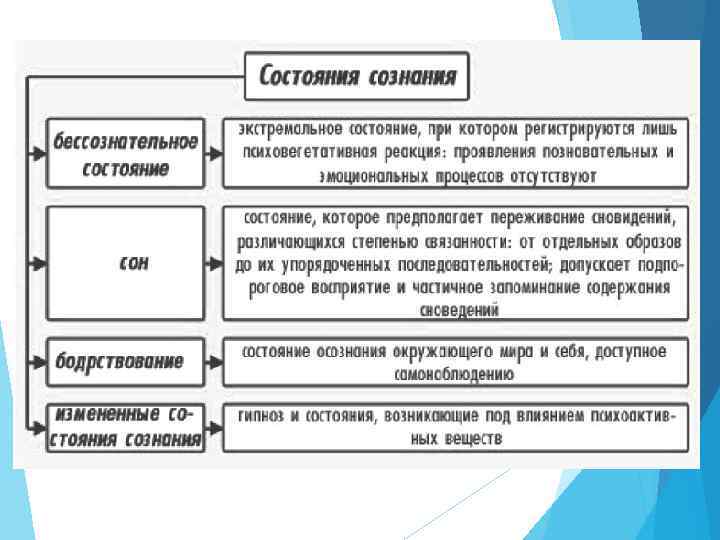

 Сон – это физиологическое состояние, при котором значительно снижается реакция нервной системы и всего организма на внешние раздражители. Состояние сна является следствием включения особых центров сна головного мозга и сопровождается снижением интенсивности физиологических процессов, общей неподвижностью.
Сон – это физиологическое состояние, при котором значительно снижается реакция нервной системы и всего организма на внешние раздражители. Состояние сна является следствием включения особых центров сна головного мозга и сопровождается снижением интенсивности физиологических процессов, общей неподвижностью.
 Выделяют две основные стадии сна - «медленный» и «быстрый» сон. Феномен «быстрого» сна открыли в 1953 г. американские ученые А. Азеринский и К. Клейтман. Медленноволновой сон человека периодически прерывается короткими периодами низкоамплитудной ЭЭГ (похожей на десинхронизацию ЭЭГ при пробуждении), сопровождающимися быстрыми движениями глазных яблок. Отсюда часто употребляемое название этой стадии - парадоксальная, или стадия быстрых движений глаз (БДГ-сон, или REM-стадия - от rapid eye movement). Люди, разбуженные во время парадоксальной стадии сна, в 80 % случаев сообщали о сновидениях. Мышцы тела расслаблены сильнее всего во время парадоксального сна, однако вегетативные показатели (дыхание, сердцебиение, кровяное давление и др.) во время парадоксального сна могут соответствовать активному бодрствованию (вплоть до так называемой «вегетативной бури»).
Выделяют две основные стадии сна - «медленный» и «быстрый» сон. Феномен «быстрого» сна открыли в 1953 г. американские ученые А. Азеринский и К. Клейтман. Медленноволновой сон человека периодически прерывается короткими периодами низкоамплитудной ЭЭГ (похожей на десинхронизацию ЭЭГ при пробуждении), сопровождающимися быстрыми движениями глазных яблок. Отсюда часто употребляемое название этой стадии - парадоксальная, или стадия быстрых движений глаз (БДГ-сон, или REM-стадия - от rapid eye movement). Люди, разбуженные во время парадоксальной стадии сна, в 80 % случаев сообщали о сновидениях. Мышцы тела расслаблены сильнее всего во время парадоксального сна, однако вегетативные показатели (дыхание, сердцебиение, кровяное давление и др.) во время парадоксального сна могут соответствовать активному бодрствованию (вплоть до так называемой «вегетативной бури»).

 График, показывающий возрастные изменения продолжительности и доли парадоксального сна у человека. Видно резкое сокращение длительности парадоксального сна (БДГ) - от 8 часов у новорожденных до 1 часа у пожилых людей. Изменения в продолжительности медленноволнового сна (МВ) не столь выражено; уменьшение от 8 часов до примерно 5 часов.
График, показывающий возрастные изменения продолжительности и доли парадоксального сна у человека. Видно резкое сокращение длительности парадоксального сна (БДГ) - от 8 часов у новорожденных до 1 часа у пожилых людей. Изменения в продолжительности медленноволнового сна (МВ) не столь выражено; уменьшение от 8 часов до примерно 5 часов.
 ЭЭГ человека при бодрствовании и сне Бодрствование Частота Амплитуда Синхронизация (регулярность колебаний) Медленноволновый сон Парадоксальный сон (БДГ) высокая низкая (нерегулярная активность, десинхронизация) высокая (синхронизированная активность) низкая (нерегулярная активность, десинхронизация)
ЭЭГ человека при бодрствовании и сне Бодрствование Частота Амплитуда Синхронизация (регулярность колебаний) Медленноволновый сон Парадоксальный сон (БДГ) высокая низкая (нерегулярная активность, десинхронизация) высокая (синхронизированная активность) низкая (нерегулярная активность, десинхронизация)
 В состоянии бодрствования с открытыми глазами преобладает бета-ритм, и этот же ритм наблюдается при парадоксальном сне. В процессе засыпания человек проходит 4 стадии, от спокойного бодрствования с закрытыми глазами (альфа-ритм) до собственно медленноволнового сна (дельта-ритм).
В состоянии бодрствования с открытыми глазами преобладает бета-ритм, и этот же ритм наблюдается при парадоксальном сне. В процессе засыпания человек проходит 4 стадии, от спокойного бодрствования с закрытыми глазами (альфа-ритм) до собственно медленноволнового сна (дельта-ритм).
 Основные ритмы ЭЭГ человека Частота Состояние 12 -25 Гц Бодрствование с открытыми глазами и БДГ-сон Альфа-ритм 8 -12 Гц Бодрствование с закрытыми глазами Тета-ритм 4 -8 Гц Засыпание 1 -4 Гц Медленноволновый сон Бета-ритм Дельта-ритм
Основные ритмы ЭЭГ человека Частота Состояние 12 -25 Гц Бодрствование с открытыми глазами и БДГ-сон Альфа-ритм 8 -12 Гц Бодрствование с закрытыми глазами Тета-ритм 4 -8 Гц Засыпание 1 -4 Гц Медленноволновый сон Бета-ритм Дельта-ритм
 ЭЭГ при различных уровнях бодрствования и сна. БДГ-сон напоминает по характеру ЭЭГ состояние бодрствования, однако электроды не регистрируют мышечной активности нигде, кроме глазных мышц.
ЭЭГ при различных уровнях бодрствования и сна. БДГ-сон напоминает по характеру ЭЭГ состояние бодрствования, однако электроды не регистрируют мышечной активности нигде, кроме глазных мышц.
 Препараты изолированного мозга (А) и изолированного переднего мозга (Б) с соответствующими записями ЭЭГ (так называемые «перерезки Бремера» - по Бремеру, 1937). При перерезке на границе головного мозга со спинным (препарат А) в ЭЭГ сохранялась картина бодрствования, а при перерезке на уровне среднего мозга (препарат Б) получали препарат спящего мозга. Бремер заключил, что в изолированном головном мозгу имеется центр пробуждения, расположенный в продолговатом и среднем мозгу.
Препараты изолированного мозга (А) и изолированного переднего мозга (Б) с соответствующими записями ЭЭГ (так называемые «перерезки Бремера» - по Бремеру, 1937). При перерезке на границе головного мозга со спинным (препарат А) в ЭЭГ сохранялась картина бодрствования, а при перерезке на уровне среднего мозга (препарат Б) получали препарат спящего мозга. Бремер заключил, что в изолированном головном мозгу имеется центр пробуждения, расположенный в продолговатом и среднем мозгу.
 Дж. Моруцци и Х. Мегун показали в 1949 г. , что стимуляция ретикулярной формации ствола мозга вызывает пробуждение. Сон при этом в то время рассматривался как следствие временной блокады активирующих восходящих влияний с одновременным «включением» таламокортикальных синхронизирующих процессов.
Дж. Моруцци и Х. Мегун показали в 1949 г. , что стимуляция ретикулярной формации ствола мозга вызывает пробуждение. Сон при этом в то время рассматривался как следствие временной блокады активирующих восходящих влияний с одновременным «включением» таламокортикальных синхронизирующих процессов.
 В настоящее время стало очевидно, что вместо единой «активирующей ретикулярной формации» (представление о которой сформировалось благодаря работам Бремера, Моруцци, Мегуна и других исследователей), существует большое количество центров, которые различаются по своим функциям, выделяемым нейромедиаторам и локализации в мозге. Большинство из них действительно находятся в ретикулярной формации ствола мозга или вблизи нее, но часть из них – в гипоталамусе и других структурах мозга.
В настоящее время стало очевидно, что вместо единой «активирующей ретикулярной формации» (представление о которой сформировалось благодаря работам Бремера, Моруцци, Мегуна и других исследователей), существует большое количество центров, которые различаются по своим функциям, выделяемым нейромедиаторам и локализации в мозге. Большинство из них действительно находятся в ретикулярной формации ствола мозга или вблизи нее, но часть из них – в гипоталамусе и других структурах мозга.
 Некоторые системы регуляции сна и бодрствования. Видно, как в цепях, выходящих из голубого пятна (медиатор - норадреналин) и ядер шва (медиатор - серотонин) аксоны расходятся к различным участкам спинного мозга, мозжечка и таламуса.
Некоторые системы регуляции сна и бодрствования. Видно, как в цепях, выходящих из голубого пятна (медиатор - норадреналин) и ядер шва (медиатор - серотонин) аксоны расходятся к различным участкам спинного мозга, мозжечка и таламуса.
 Выделение модулирующих медиаторов в кору больших полушарий во время бодрствования и сна Бодрствование Медленноволновый сон Парадоксальный сон Преобладающий ритм в ЭЭГ бета-ритм дельта-ритм бета-ритм Выделение норадреналина (из голубого пятна) максимально снижено отсутствует Выделение серотонина (из дорзальных ядер шва) максимально снижено отсутствует Выделение гистамина (из туберомамиллярного ядра заднего гипоталамуса) максимально снижено отсутствует Выделение ацетилхолина (из базальных ядер переднего мозга - базальное крупноклеточное ядро и др.) максимально отсутствует максимально
Выделение модулирующих медиаторов в кору больших полушарий во время бодрствования и сна Бодрствование Медленноволновый сон Парадоксальный сон Преобладающий ритм в ЭЭГ бета-ритм дельта-ритм бета-ритм Выделение норадреналина (из голубого пятна) максимально снижено отсутствует Выделение серотонина (из дорзальных ядер шва) максимально снижено отсутствует Выделение гистамина (из туберомамиллярного ядра заднего гипоталамуса) максимально снижено отсутствует Выделение ацетилхолина (из базальных ядер переднего мозга - базальное крупноклеточное ядро и др.) максимально отсутствует максимально
 В настоящее время очевидно, что бодрствование и сон – качественно различные состояния, каждое из которых имеет свои управляющие центры.
В настоящее время очевидно, что бодрствование и сон – качественно различные состояния, каждое из которых имеет свои управляющие центры.
 Предполагаемые центры бодрствования: 1 - базальные ядра переднего мозга (выделяют ацетилхолин), 2 - ядра заднего гипоталамуса (туберомаммиллярное ядро) (выделяют гистамин), 3 - дорзальные ядра шва (выделяют серотонин), 4 - область покрышки моста (выделяет ацетилхолин и глутамат), 5 - голубое пятно (выделяет норадреналин). Схема расположения «центров бодрствования» в головном мозге человека и влияния на них орексиновых нейронов (показано красным). Орексиновые нейроны немногочисленны и расположены в гипоталамусе Предполагается, что именно выделение нейропептида орексина нейронами этих ядер переключает мозг в режим бодрствования.
Предполагаемые центры бодрствования: 1 - базальные ядра переднего мозга (выделяют ацетилхолин), 2 - ядра заднего гипоталамуса (туберомаммиллярное ядро) (выделяют гистамин), 3 - дорзальные ядра шва (выделяют серотонин), 4 - область покрышки моста (выделяет ацетилхолин и глутамат), 5 - голубое пятно (выделяет норадреналин). Схема расположения «центров бодрствования» в головном мозге человека и влияния на них орексиновых нейронов (показано красным). Орексиновые нейроны немногочисленны и расположены в гипоталамусе Предполагается, что именно выделение нейропептида орексина нейронами этих ядер переключает мозг в режим бодрствования.
 Функция орексина как медиатора, ответственного за координацию других центров бодрствования, была открыта совсем недавно – около 2003 г. , а сам орексин был открыт лишь в 1998 г. При потере хотя бы части орексиновых нейронов возникает нарколепсия – дневная сонливость.
Функция орексина как медиатора, ответственного за координацию других центров бодрствования, была открыта совсем недавно – около 2003 г. , а сам орексин был открыт лишь в 1998 г. При потере хотя бы части орексиновых нейронов возникает нарколепсия – дневная сонливость.
 Схема центра расположения центра медленного сна. Вверху – бодрствование, когда центр сна заторможен и ретикулярная формация (РФ) активирует кору; внизу – центр сна возбужден, РФ заторможена и кора не активируется.
Схема центра расположения центра медленного сна. Вверху – бодрствование, когда центр сна заторможен и ретикулярная формация (РФ) активирует кору; внизу – центр сна возбужден, РФ заторможена и кора не активируется.
 Предполагаемые центры сна: Центр медленноволнового сна: в переднем гипоталамусе, (в вентролатеральном преоптическом ядре), медиатор - гаммааминомасляная кислота (ГАМК); активность нейронов незначительна в бодрствовании, но резко возрастает в период обычного сна и прекращается во время парадоксального. Центр парадоксального сна: ретикулярная формация покрышки моста (латеродорзальная / педункуло-понтийная области покрышки моста), медиаторы - ацетилхолин и глутамат.
Предполагаемые центры сна: Центр медленноволнового сна: в переднем гипоталамусе, (в вентролатеральном преоптическом ядре), медиатор - гаммааминомасляная кислота (ГАМК); активность нейронов незначительна в бодрствовании, но резко возрастает в период обычного сна и прекращается во время парадоксального. Центр парадоксального сна: ретикулярная формация покрышки моста (латеродорзальная / педункуло-понтийная области покрышки моста), медиаторы - ацетилхолин и глутамат.
 Список Литературы Дубынин В. А. Регуляторные системы организма человека. М. : Дрофа, 2003. Дудьев В. П. Психомоторика: cловарь-справочник, 2008 г. Мэгун Г. Бодрствующий мозг. М. : Мир, 1965. Росси Я. А. , Цанкетти А. Ретикулярная формация ствола мозга. М. : ИЛ, 1960. Шульговский В. В. Физиология центральной нервной системы. М. : Изд-во Моск. ун-та, 1987.
Список Литературы Дубынин В. А. Регуляторные системы организма человека. М. : Дрофа, 2003. Дудьев В. П. Психомоторика: cловарь-справочник, 2008 г. Мэгун Г. Бодрствующий мозг. М. : Мир, 1965. Росси Я. А. , Цанкетти А. Ретикулярная формация ствола мозга. М. : ИЛ, 1960. Шульговский В. В. Физиология центральной нервной системы. М. : Изд-во Моск. ун-та, 1987.
К настоящему времени оформились представления о существовании в мозгу двух систем, регулирующих сон и бодрствование.
Одна из них - восходящая активирующая ретикулярная система - расположена в верхних отделах ретикулярной формации ствола мозга и задних отделах гипоталамуса. При раздражении этой системы на электроэнцефалограмме появляется десинхронизация, уплощение и ускорение ритмов, что у спящих животных сопровождается пробуждением, а у бодрствующих повышением бдительности. Все периферические стимулы оказывают влияние на эту систему через коллатерали, которые отходят к ней от чувствительных путей, идущих к коре больших полушарий. Непосредственное электрическое раздражение корковых полей и некоторых других глубинных образований мозга может также вызвать пробуждение.
Но теперь уже очевидно, что раздражение всех отделов мозга, а также активность систем мозга, воспринимающих внешние и внутренние влияния, оказывают пробуждающее воздействие через восходящую активирующую систему. Опыты с перерезкой мозга и повреждением верхних отделов ретикулярной формации подтверждают это положение: животные погружаются в сонное состояние, из которого вывести их не удается.
Недавно появились сообщения, что при хорошем уходе у животных после операции через несколько недель появляются признаки бодрствования, увеличивающиеся со временем. Что это значит? Имеется ли еще одно звено активирующей системы, которое берет на себя осуществление этой функции, или в описанных опытах разрушается не вся восходящая система, сказать трудно. Возможным представляется существование активирующих аппаратов в лимбических структурах (миндалина, гиппокамп, таламус), функционально тесно связанных с аппаратами ретикулярной формации и гипоталамуса.
Более сложно построена вторая система - гипногенная, активность которой определяет длительность и глубину сна.
К настоящему времени уточнена роль ряда структур головного мозга в организации сна. Начнем с нижних отделов ствола. Моруцци описал синхронизирующий аппарат, при раздражении которого возникают электрофизиологические и поведенческие проявления сна. Роль этого образования сейчас хорошо выявлена: при отделении его (путем перезрезки) продолжительность сна у кошки уменьшается более чем в три раза. Животное бодрствует большую часть суток.
Разработан интересный способ анализа: в артерию вводят наркотическое вещество, выключающее временно функции определенных структур. Введение наркотика в сосуд, снабжающий нижний ствол кровью, приводит к тем же результатам, что и перерезка: удлиняется время бодрствования.
Этот аппарат тесно связан с каротидным синусом - образованием, расположенным в развилке наружной и внутренней сонных артерий, которое сигнализирует в мозг об уровне артериального давления и некоторых химических показателей. Раздражение каротидного синуса ведет к усилению деятельности синхронизирующего заднестволового аппарата, снятие раздражения - к обратному эффекту.
Роль барорецепторов этой зоны была подмечена уже давно, ведь не случайно артерии носят название «сонных». Известно, что в Индонезии на острове Бали знахари двухминутным массажем каротидного синуса вызывают сон. Совсем недавно французские нейрофизиологи описали в области нижнего ствола еще один синхронизирующий аппарат.
Другая гипногенная зона находится в области переднего гипоталамуса и перегородки. Раздражение этих структур электрическим током любой частоты приводит к синхронизации электроэнцефалографических ритмов и наступлению сна. Животное проделывает все ритуалы, характерные для его естественного сна (облизывание, мышечное расслабление, зевота). Разрушение этого аппарата приводит к длительному бодрствованию и резким нарушениям восстановительных процессов.
Еще одно важное звено в системе синхронизирующих аппаратов - таламическая синхронизирующая система. Раздражение низкочастотным электрическим током определенных ядер таламуса приводит к синхронизации потенциалов мозга и сну. Некоторые исследователи считают его главной гипногенной структурой, так как сон, наступающий при его раздражении, длителен и неотличим от нормального, а также вызывается легче, чем при раздражении других структур.
При низкочастотном раздражении сон можно вызвать, воздействуя на другие структуры мозга и даже периферические нервы. (Высокочастотное раздражение, как правило, приводит к пробуждению и десинхронизации.) Все это свидетельствует о распространенности синхронизирующих и десинхронизирующих аппаратов в нервной системе. Несомненно имеются сгущения, где они представлены более значительно. При разрушении этих скоплений и возникают эффекты противоположного характера - уменьшение или увеличение длительности сна.
Таким образом, имеются три главные гипногенные зоны, обеспечивающие возникновение и развитие сна. Мы знакомы с двумя видами сна, поэтому необходимо подчеркнуть, что эти структуры обеспечивают медленный сон. Как уже говорилось, за быстрый сон ответственны структуры средних отделов ствола мозга (ретикулярные ядра Варолиева моста). При их разрушении быстрый сон не наступает.
Гипногенная система по своей архитектуре сложна и включает многие аппараты мозга. Химически она, вероятно, неоднородна, так как в качестве медиаторов используются ацетилхолин, серотонин, гамааминомасляная кислота - ГАМК.
Что же представляет собой сон по своим физиологическим механизмам? Сразу же отпадает точка зрения, согласно которой сон - это отсутствие бодрствования, т. е. в основе его лежит выключение активирующих аппаратов. Очевидно, что имеются механизмы, организующие сон. Главное же заключается в том, что сон является активным организованным процессом, включающим различные по своей сущности и физиологическим механизмам состояния. Вот почему так сложно организована гипногенная система. Сон - сочетание активного состояния специализированных синхронизирующих аппаратов и снижения активности активирующей восходящей системы. Данные о состоянии отдельных нейронов во время сна хорошо подтверждают это положение. Следовательно, сами по себе отпадают представления о сне как охранительном, разлитом торможении. Только внешне это состояние можно охарактеризовать так. Однако приглядевшись, и в состоянии скелетно-мышечной системы можно видеть активность. Интенсивная психическая деятельность во время сна также говорит об активности мозга в этом состоянии.
Итак, существуют две системы, регулирующие сон и бодрствование. У систем имеются подсистемы, включающие различные формы сна в определенной последовательности. Все наводит на мысль о существовании в мозгу координирующего аппарата, который в определенное время регулирует включение отдельных систем в целом, а затем и их подсистем. В этом нас убеждают наблюдения над больными людьми, когда все подсистемы работают, но резко нарушается закономерная последовательность их включения. Координирующий аппарат не находится в одном каком-либо отделе мозга. Речь идет о сложном комплексе с преимущественным расположением в передних отделах больших полушарий мозга, лимбических аппаратах, гипоталамусе. Дальнейшие исследования позволят более четко и обоснованно представлять подобную точку зрения.
Освещая современные представления о регуляции сна и бодрствования, нельзя не вернуться к гуморальным факторам в происхождении сна. Поиски гипнотоксинов - веществ, накопление которых вызывает сон, ведутся очень давно. Имеется целый ряд исследований, результаты которых трудно объяснить без участия какого-то гуморального агента. В исследовании немецкого физиолога Кроля показано, что экстракт вещества головного мозга спящего животного при внутривенном введении его вызывает сон у подопытного животного. Выше уже были описаны опыты (Монье, Корнмюллера): у подопытного животного наступал сон, если в его организм поступала кровь из мозга другого животного, погруженного в сон в результате раздражения зрительного бугра.
Известным физиологом А.В. Тонких была показана роль гормонов, главным образом гипофиза, в возникновении сна. В Лаборатории по изучению нервных и гуморальных регуляций имени Н.И. Гращенкова также проведены специальные исследования содержания активных биологических веществ в крови и моче у больных с повышенной сонливостью. Было установлено, что содержание в крови и в моче адреналина (гормонов коркового слоя надпочечника) снижено, а ацетилхолина, гистамина и продуктов обмена серотонина повышено.
Эти данные несомненно интересны, однако возникает вопрос, насколько содержание биологически активных веществ на периферии отражает их истинные соотношения в головном мозге. На примере сиамских близнецов опровергается гуморальная теория возникновения сна. Следует иметь в виду, что в циркулирующих жидкостях содержание активных биологических веществ одинаково, а в мозгу различно. Возможно, именно поэтому одна голова спала, а другая в это время бодрствовала. Вот почему в последние годы стали уделять особое внимание химическим передатчикам нервных импульсов, богато представленным в мозге.
В настоящее время очевидно присутствие в мозге медиаторов - веществ, выделяющихся в синапсах на границе двух нейронов и обеспечивающих распространение нервного импульса. Холинэргические синапсы в качестве медиатора выделяют ацетилхолин, адренэргические - норадреналин, серотонинэргические - серотонин. Есть синапсы с гамма-аминомасляной кислотой и большое число синапсов с еще не идентифицированным химическим передатчиком. Все химически неоднородные нейроны не разбросаны в мозгу хаотично, а составляют определенные системы, объединяемые по принципу представленности в них того или иного медиатора. Норадреналин и серотонин обнаруживаются главным образом в глубинных и стволовых структурах мозга, в то время как ацетилхолин распределен более равномерно.
Различным химическим системам придавалось и определенное функциональное значение. На опытах доказано, что активирующая восходящая ретикулярная система, поддерживающая необходимый уровень бодрствования, по своей химической характеристике адренэргическая, что введение адреналина усиливает настороженность животного, а во сне содержание его в мозгу снижается. Многие фармакологические средства, препятствующие сну, близки по составу к адреналину либо, вмешиваясь в химию мозга, способствуют накоплению этих веществ. Правда, установлено, что нередко одна функциональная система является гетерохимической, т. е. в нее входят нейроны, медиаторы, которые различны по химическому составу.
В последнее время формируется представление, согласно которому основные гипногенные вещества - ацетилхолин, серотонин и ГАМК. Физиолог из Южной Америки Эрнандец-Пеон обнаружил с помощью специальных опытов, что наложение кристаллика ацетилхолина на структуры ствола мозга, гипоталамуса, медиальных отделов височной доли вызывает электроэнцефалографические и поведенческие признаки сна. Косвенным доказательством является и то, что в отделах мозга, где расположены гипногенные аппараты, основной медиатор - ацетилхолин.
Накапливаются также факты, которые говорят о роли серотонина. Разрушения ядер шва, расположенных в стволе мозга и наиболее богатых серотонином, приводят к бессоннице, степень которой обратно пропорциональна числу сохраненных ядер. Проведены многочисленные опыты с введением в организм аминокислоты - предшественника серотонина - триптофана и антагонистов серотонина - метисергида, дезерила, разрушающих его и оказывающих противоположное действие на сон в целом и на его отдельные фазы. По этому вопросу возникла дискуссия: одни ученые отстаивают мнение, согласно которому серотонин способствует возникновению быстрого сна, другие - медленного. По-видимому, более обоснованна вторая точка зрения. Удалось показать, что адренэргические аппараты участвуют не только в механизмах бодрствования, но и быстрого сна. Эти исследования имеют огромное практическое значение, так как являются базой для создания современной дифференцированной фармакологии сна и бодрствования.
Перспективы этой проблемы заключаются скорее всего не в поисках каких-то особых гипногенных веществ, а в выяснении истинной роли уже известных химических активных агентов и в идентификации еще не опознанных медиаторов головного мозга.
Интерес к гуморальным исследованиям особенно обострился в связи с открытием быстрого сна. Было установлено, что лишение людей и животных быстрого сна приводит к увеличению этой фазы сна в последующие ночи. Создалось впечатление, что в фазе быстрого сна разрушается какое-то гипотетическое вещество, накапливающееся во время бодрствования. Следовательно, при лишении быстрого сна этот фактор продолжает накапливаться и в последующие ночи вызывает избыточную длительность этой фазы. Однако против подобной гипотезы говорит ежедневное наблюдение, связанное с большей продолжительностью быстрого сна во второй половине ночи, когда накопившееся вещество должно было бы уже разрушиться. Тем не менее такими общими, хотя и логическими соображениями противостоять указанной гипотезе трудно. Нужны факты. Пока же создается впечатление, что отдельные фазы сна имеют собственную химию.
В основе регуляции смены активного и неактивного состояния животного организма лежат нервные и гуморальные процессы.
Для многих позвоночных животных определяющее значение в возникновении ритмов поведения имеет свет. Прежде всего это относится к птицам. У подавляющего большинства пернатых (исключение составляют ночные виды) с приближением вечерних сумерек развивается сонливость, дремота и сон. Утром на рассвете они просыпаются и включаются в активные действия. Такой же биоритм активности характерен и для млекопитающих животных с монофазной природой сна. У животных с полифазной природой сна связь с фотопериодом менее выражена или вообще отсутствует.
Усилиями физиологов, морфологов, биохимиков и гистохимиков доказано, что нервные клетки ядер шва обладают ритмической нейросекрецией. Снижение интенсивности афферентного потока со стороны экстеро-, интеро — и проприорецепторного аппарата организма животных приводит к падению неспецифической электрической активности ретикулярной формации ствола мозга, стимулирует функциональную активность ядер шва и позволяет неспецифическим ядрам таламуса синхронизировать электрическую активность коры.
Стимуляция ядер шва, в свою очередь, запускает процесс синтеза серотонина из аминокислоты триптофана. Образующийся в ядрах шва серотонин по аксонам нейронов распространяется к нейронам ретикулярной формации, таламуса, гипоталамуса, лимбической системы и блокирует их активирующее влияние на кору больших полушарий. Отсюда следует справедливое утверждение о том, что серотонин - это не просто медиатор центральной нервной системы, а медиатор сна . Однако его участие в механизме формирования сонного состояния специфично.
В специальных экспериментах было обнаружено, что искусственная блокада процессов синтеза серотонина устраняет у животных лишь фазу медленного сна и не влияет на фазу быстрого сна. Таким образом, серотонин выступает как посредник медленного сна .
Нейросекреторной активностью обладает еще одна область ствола мозга - так называемое синее пятно покрышки. Здесь вырабатывается норадреналин - медиатор пробуждения . Он выступает в качестве антагониста серотонина. Активность синего пятна, кроме того, приводит к торможению функциональной активности ядер шва. Другими словами, синее пятно покрышки и ядра шва находятся в реципрокных отношениях.
Интересную гипотезу регуляции ритма сна и бодрствования предложили американские исследователи А. Хобсон и Р. Мак-Карли (1977). В соответствии с их представлениями, биоритмы сна задаются спонтанной электрической активностью гигантских нейронов моста, которые имеют синаптические связи со многими структурами головного мозга. Ритмическая электрическая активность гигантоклеточного ядра моста, адресованная синему пятну, служит триггерным механизмом пробуждения. Активность гигантских нейронов моста, направленная на ядра шва, приводит к интеграции тормозных процессов и развитию сна. В данной схеме остаются неясными причины активизации тех или иных гигантских нейронов.
Условием жизни человека является периодическая смена сна и бодрствования. Сон является разновидностью внутреннего торможения. И.П. Павлов рассматривал сон как внутреннее торможение, распространившееся на всю массу полушарий и нижележащие отделы мозга (до продолговатого мозга).
Сон – периодическое функциональное состояние человека, характеризующееся отсутствием целенаправленной деятельности и активных связей с окружающей средой. Во время сна активность мозга не уменьшается, а перестраивается.
Сон является нормальной потребностью человека, такой же, как в пище и воде. Лишь в течение 4 – 5 дней человек может преодолевать сон, по истечении которых он засыпает в любой позе. Так, описаны случаи, когда наездник засыпал, сидя в седле, на полном ходу лошади.
Современные научные данные говорят о том, что полностью работа мозга во сне не прекращается, происходит лишь видоизменение жизни мозга. Приборами зафиксировано, что биотоки коры во время сна изменяют свой ритм, но совсем не исчезают. В отдельных случаях целые зоны мозга продолжают напряженную дневную работу. Этим объясняется, что во время сна у Д.И. Менделеева окончательно сложилась периодическая система элементов. Но такие случаи являются исключительными и, как правило, во сне человек мыслит хаотично, отдельными беспорядочными образами.
С помощью электрофизиологических исследований удалось обнаружить нервные структуры, принимающие участие в регуляции состояния сна и бодрствования, расположенные в подкорковых отделах.
Наиболее важная роль принадлежит таламусу, гипоталамусу и ретикулярной формации.
В таламусе и гипоталамусе обнаружены центры сна (их раздражение с помощью вживленных электродов вызывало сон у животных). В ретикулярной формации – центры, раздражение которых приводило к активации мозговой деятельности и поддерживало бодрое состояние. В последние десятилетия была доказана и роль коры головного мозга в регуляции собственного уровня функциональной активности.
При регистрации биотоков мозга во время сна было установлено, что сон проходит в несколько стадий.
Во время глубокого снанаблюдается высокоамплитудный медленный дельта-ритм с частотой 4-5 колебаний в секунду и менее. Такой сон называют медленным или ортодоксальным.
Медленный сон (ортодоксальный, классический) наступает после засыпания, длится 60-90 минут. Снижается обмен веществ и активность сердечно-сосудистой, дыхательной, пищеварительной и выделительной систем, мышцы расслабляются, температура понижается. Психические процессы мозга: сновидения почти отсутствуют. Может происходить разговор во сне, возникают ночные страхи у детей и снохождение (лунатизм).
Быстрый сон (парадоксальный)
был открыт в 1953 г. Э. Азеринским и Н.
Клейтменом. Он наступает после медленного сна, длится 10 – 15 минут. В электроэнцефалограмме появляются быстрые высокочастотные ритмы, похожие на ритмы бодрствующего человека. Активизируется деятельность внутренних органов: учащается пульс, дыхание, повышается температура, сокращаются глазодвигательные мышцы (глазные яблоки двигаются), мимические мышцы и мышцы конечностей. Сон не глубокий и поверхностный, но разбудить человека в это время довольно сложно (поэтому парадоксальный).
Психические процессы мозга : наблюдается возбуждение нейронов затылочных долей, происходит классификация и упорядочение поступившей за день информации, появляются реалистические эмоциональные сновидения со зрительными, звуковыми и обонятельными образами. Если человека разбудить, то он всегда может рассказать о своих сновидениях (во время медленного сна сновидения не помнят). Чаще всего мы просыпаемся именно в эту стадию, поэтому помним сны.
Важное влияние на сон могут оказывать внешние и внутренние раздражители, которые бессознательно регистрируются мозгом и включаются в сюжет сновидений. Например, раскаты грома на улице, а человеку снится пушечная стрельба; переполненный желудок может вызвать чувство удушения и т. д.
Лишение человека «парадоксального сна» может привести к расстройствам памяти и психическим заболеваниям. У человека при лишении сна в течении 3 – 5 суток возникают утомление, галлюцинации, функции ВНД упрощаются. На 7 сутки наблюдается нарушение сознания. В опытах на животных собаки погибали на 17 – 21 день лишения сна. При этом обнаруживались морфологические изменения в ЦНС.
За время ночного сна у человека наблюдается 3 – 5 периодических смен медленного и быстрого сна. За ночь человек видит 5 – 6 снов. У взрослых стадия быстрого сна составляет примерно 25 % от общего времени сна, а у детей – значительно больше.
Продолжительность сна зависит от возраста:
Новорожденный 21 час
6 мес – 1 год 15 часов
4 года 12 часов
10 лет 10 часов
14 лет 8 – 9 часов
взрослые 7 – 8 часов
Человек проводит во сне третью часть жизни: из 75 лет – 25 лет спит. Сокращение времени сна способствует ухудшению общего самочувствия, снижает физическую и умственную работоспособность.
Засыпанию могут мешать:
1) внешние сигналы (яркий свет, громкий звук, духота в помещении);
2) внутренние сигналы (боль, чувство голода, переполненный желудок, беспокойные мысли);
3) гуморальные сигналы (адреналин).
Способствуют засыпанию:
1) приспособленность к смене дня и ночи;
2) условные рефлексы на привычную обстановку спальни, время сна;
3) внешнее воздействие (монотонность, тепло);
4) внутреннее воздействие (усталость, скука);
5) гуморальное воздействие (серотонин).
Это сложный физиологический процесс, имеющий локализованный анатомический субстрат (ретикулярная субстанция, кора больших полушарий, область гиппокампа, поясная извилина, таламус). Механизмы циркадного и ультрадианного ритмов - это важный компонент всей рерулирующей системы.
В регуляцию цикла сон\бодрствование осуществляется с помощью следующих механизмов:
1) Механизмы внутрисуточного ритма активность-покой.
2) Механизмы восходящей активации (бодрствование).
3) Механизмы обычного (медленного) сна.
4) Механизмы парадоксального сна.
Двухкомпонентная модель регуляции сна.
1) Гомеостатический фактор (фактор S).
Δ-индекс в момент пробуждения претерпевает нарастание.
2) Циркадный фактор (фактор C).
Определяется по характеру температуры тела. Когда фактор накапливается, человек засыпает, и лишь физическая нагрузка может заставить его бодрствовать некоторое время.
Наше состояние в данный момент определяется алгебраической суммой двух кривых. В каждом эпизоде сна интенсивность медленноволновой фазы (светлая штриховка) постепенно снижается, а быстрой (парадоксальной) – возрастает. W – бодрствование, S – сон.
Третий фактор - фактор U – это ультрадианный сон – ночной сон
, во время которого происходит чередование симпатической и парасимпатической регуляции.
Таким образом, три основных процесса вовлеченны в регуляцию сна. Длительность и интенсивность сна поддерживается гомеостазом, циркадный ритм определяет периоды нарастания «давления» сна, а ультрадианные механизмы лежат в основе циклического чередования медленного и быстрого сна.
Под циркадными ритмами
подразумевают физиологические и психологические явления, периодически возникающие в течение суток. Большинство циркадианных ритмов регулируется супрахиазмальными ядрами переднего гипоталамуса. Что касается подготовки ко сну, то за час до наступления сна при неярком освещении происходит выброс мелатонина
. Следует отметить, что взаимодействие супрахиазмальных ядер и эпифиза, вырабатывающего мелатонин, реципроктно.
Механизмы бодрствования, медленного сна.
Во время бодрствования
происходит тоническая деполяризация коры головного мозга
.
Существует несколько систем, регулирующих бодрствование. Это:
1) Система голубого пятна (медиатор – норадреналин).
2) Педункулопонтийная область моста (медиатор – ацетилхолин).
3) Ядра шва (медиатор – серотонин).
4) Мезэнцефалическая ретикулярная формация (медиатор – глутамат).
5) Вентральная тегментальная область, а также компактная часть черного вещества (медиатор – дофамин).
6) Задний латеральный таламус (пептид гипокритий – орексин).
7) Туберо-маммилярное ядро (медиатор – гистамин).
8) Базальная часть переднего мозга (медиатор – ацетилхолин).
9) Супрахиазмальные ядра.
10) Медиальная префронтальная кора.
Системы восходящей активации.
1. Классический путь: из таламических ядер глутамат поступает в кору.
2. Вентральный путь: по медиальному пучку переднего мозга, из базальные ядер в кору поступают холинергические импульсы.
Система медленного сна.
Во время медленного сна кора находится в состоянии тонической гиперполяризации
. Первично происходит ослабление центров, отвечающих за бодрствование. Далее включается тормозная система и усиливает торможение этих центров. Во время сна нейроны работают в режиме пачка- пауза.
Системы торможения:
1) Передний таламус, преоптическое ядро, в которых находятся ГАМК-нейроны. Стоит отметить, что клетки этой области имеют длинные аксоны, в везикулах которых находится пептид галантин; он придает импульсам дополнительную силу и продолжительность.
2) Короткоаксонные холинергические клетки в I-II слоях коры головного мозга
3) Торможение происходит также при подаче импульсов на пирамидальный корково-таламический нейрон, во время чего возникают пучки деполяризующих сигналов на таламус.
Несмотря на глубокое торможение, передача информации не прекращается
: наблюдаются кальциевые спайки на фоне гиперполяризации (колебания мембраны).
Во время медленного сна головной мозг восстанавливает метаболический баланс, баланс электролитов
. Субъективное ощущение выспанности наступает восле восстановленияитих параметров, то есть через 2-4 цикла сна.
